Минеральные вещества входят в состав всех живых тканей. Однако нормальное функционирование тканей обеспечивается не только наличием в них тех или иных минеральных солей, но и строго определенным их соотношением. Минеральные вещества поддерживают необходимое осмотическое давление в биологических жидкостях и обеспечивают постоянство кислотно-щелочного равновесия в организме.Рассмотрим основные минеральные вещества.
Калий содержится главным образом в клетках, натрий - в межклеточной жидкости. Для нормальной жизнедеятельности организма требуется строго определенное соотношение частиц натрия и калия. Должное соотношение этих ионов обеспечивает нормальную возбудимость нервной и мышечной тканей. Натрий играет большую роль в поддержании постоянства осмотического давления. При пониженном содержании калия в миокарде (мышечной ткани сердца) нарушается сократительная функция сердца. Но при избытке калия деятельность сердца также нарушается. Суточная потребность взрослого человека: натрий - 4-6 г, калий - 2-3 г.
Кальций входит в состав костей в виде фосфорных солей. Его ионы обеспечивают нормальную деятельность мозга и скелетных мышц. Наличие кальция необходимо для свертывания крови. Избыток кальция повышает частоту и силу сердечных сокращений, а при сверхбольших концентрациях в организме может вызвать остановку сердца. Суточная потребность взрослого человека в кальции - 0,7-0,8 г.
Фосфор входит в состав всех клеток и межтканевых жидкостей. Он играет большую роль в обмене белков, жиров, углеводов и витаминов. Это вещество - непременная составляющая богатых энергией веществ. Соли фосфорных кислот поддерживают постоянство кислотно-щелочного равновесия крови и других тканей. Суточная потребность взрослого человека в фосфоре - 1,5-2 г.
Хлор содержится в организме главным образом в соединении с натрием и входит в состав соляной кислоты желудочного сока. Хлор необходим для нормальной жизнедеятельности клеток. Суточная потребность взрослого человека в хлоре - 2-4 г.
Железо является составной частью гемоглобина и некоторых ферментов. Обеспечивая транспорт кислорода, оно принимает участие в окислительных процессах. Суточная потребность в железе для мужчин составляет 10 мг, для женщин - 18 мг.
Бром в небольших количествах содержится в крови и в других тканях. Усиливая торможение в коре больших полушарий, он способствует нормальному соотношению между процессами возбуждения и торможения.
Йод - обязательный компонент гормона щитовидной железы. Недостаток этого вещества в организме вызывает нарушение многих функций. Суточная потребность в йоде для взрослых здоровых людей составляет 0,15 мг (150 мкг).
Сера входит в состав многих белков. Она содержится в некоторых ферментах, гормонах, витаминах и других соединениях, играющих важную роль в обмене веществ. Кроме того, серная кислота используется печенью для нейтрализации некоторых веществ.
Для нормальной жизнедеятельности организма, кроме перечисленных веществ, имеют значение магний, цинк и т. д. Некоторые из них (алюминий, кобальт, марганец и др.) входят в состав организма в столь незначительных количествах, что их называют микроэлементами. Разнообразное питание обычно полностью обеспечивает организм всеми минералами.
Длинные тонкие мышечные волокна, из которых построена скелетная мышца, – это гигантские клетки, образующиеся в ходе онтогенеза при слиянии множества отдельных клеток. У взрослого человека они могут достигать в длину 5 см! Многочисленные ядра в такой клетке располагаются прямо под цитоплазматической мембраной, а основная часть цитоплазмы состоит из вытянутых вдоль всей клетки миофибрилл (толщиной 1–2 мкм), с характерной поперечной исчерченностью (рис. 3). Такую «раскраску» миофибрилле придают саркомеры, в состав каждого из которых входит два набора параллельных, частично перекрывающихся филаментов: толстых миозиновых, которые образуют темную полосу и тянутся от одного края до другого, и тонких актиновых, лежащих в области светлой полосы и частично заходящих в область темных полос (рис. 4). Один саркомер от другого отделяется Z-диском.
Рис. 3. Схема небольшого отрезка клетки скелетной мышцы (мышечного волокна)
Рис. 4. А. Электронная микрофотография продольного среза через клетку скелетной мышцы кролика (при малом увеличении). Видна регулярная поперечная исчерченность. Клетка содержит множество параллельных миофибрилл (см. рис. 3). Б. Небольшой участок того же фото: показаны отрезки двух смежных миофибрилл и детали саркомера. В. Схема строения отдельного саркомера, объясняющая происхождение темных и светлых полос, которые видны на электронной микрофотографии
Цитоплазма обеспечивает миофибриллы энергией в виде АТФ – в активно функционирующей мышце обнаруживается много митохондрий. Кроме того, в цитоплазме содержатся гликоген, фосфокреатин, гликолитические ферменты.
Скелетная мышца превращает химическую энергию, накопленную в АТФ, в механическую с очень большой эффективностью – в виде тепла теряется всего 30–50%. (Для сравнения: автомобильный двигатель при сжигании бензина обычно теряет в виде тепла 80–90%.)
При мышечном сокращении за счет связывания миозином актина толстые и тонкие нити скользят относительно друг друга (рис. 5).

Рис. 5. Схема, иллюстрирующая процесс мышечного сокращения по принципу скользящих нитей: толстые и тонкие филаменты скользят друг по другу, не изменяя собственной длины
Физиологическим регулятором сокращения мышц служат ионы кальция. В состоянии покоя работает система активного транспорта ионов кальция, и они накапливаются в своеобразном хранилище, из которого высвобождаются под действием нервного импульса, обеспечивая мышечное сокращение.
Система транспорта ионов кальция работает за счет энергии АТФ. Того количества АТФ, которое имеется в мышце, хватает на поддержание работы сократительного аппарата всего в течение доли секунды. Как же работает мышца более продолжительное время? Оказывается, в мышце энергия запасается в форме фосфокреатина, или креатинфосфата, который может переносить больше высокоэнергетических фосфатных групп, чем универсальный АТФ. Фосфокреатин восстанавливает АТФ, обеспечивая тем самым приток энергии для мышечного сокращения. Однако в работающей мышце запасы фосфокреатина быстро истощаются, а это снижает и содержание АТФ.
При более продолжительной физической нагрузке мышцы обеспечиваются энергией за счет гликолиза – расщепления углеводов под действием ферментов с накоплением энергии в виде АТФ. Когда запасы креатина в мышце истощаются, понижается энергетический заряд мышечного сокращения, что приводит к стимуляции гликолиза, цикла трикарбоновых кислот и окислительного фосфорилирования в работающей мышце.
В отсутствие кислорода при расщеплении молекулы углеводов образуются две молекулы молочной кислоты (или лактата) и две молекулы АТФ. Однако, если для гликолиза используется гликоген мышц, то возникают две молекулы лактата и три молекулы АТФ.
Гликоген – это, как известно, главный резервный полисахарид, запасаемый в мышцах и печени. При пониженном уровне гликогена в мышцах и печени и наличии свободной глюкозы в крови она используется для синтеза гликогена. И, наоборот, при потребностях организма в энергетическом источнике для процессов гликолиза успешно используется гликоген.
Мышечная система наиболее развита по сравнению с другими системами организма. Для обеспечения работы мышц необходимо огромное количество энергии, которую человек может получать за счет креатинфосфата (фосфокреатина), углеводов в виде гликогена и глюкозы и жиров. Эти три вида энергоносителей различаются между собой по величине освобождаемой при их использовании энергии и по тому, как долго может каждый из них служить «топливным» источником. Так устроен механизм обеспечения мышц энергией (табл. 2, 3). Хорошо известно, что при продолжительной неинтенсивной работе при протекании окислительных процессов используются жиры или углеводы, а при работе несколько большей интенсивности используются механизмы анаэробного гликолиза. При очень интенсивной кратковременной нагрузке работа мышц обеспечивается за счет фосфагенов. Соответственно каждый из источников энергии имеет свою энергетическую стоимость и используется при определенных условиях (табл. 4).
Работающие мышцы для аэробного окисления углеводов по сравнению с другими органами потребляют очень большое количество кислорода (табл. 5).
Таблица 2. Запасы энергии в организме человека массой тела 70 кг
Таблица 3. Максимально возможная мощность скелетных мышц человека при использовании различных субстратов и путей катаболизма
Таблица 4. Максимальная скорость образования энергии из различных источников во время физических упражнений
Источник энергии |
Максимальная скорость мышцы |
Количество в мышце, ммоль/кг образования богатых энергией фосфатных связей, ммоль/ сек/кг |
Максимальная скорость продукции, ккал/ч/кг |
Время поддержания максимальной скорости |
Креатинфосфат |
||||
Анаэробный гликолиз |
||||
Аэробное окисление глюкозы и гликогена |
||||
Аэробное окисление |
Не лимитировано |
Не лимитировано |
Таблица 5. Относительное потребление кислорода различными органами человека в покое и при тяжелой работе*
Тонус скелетных мышц. В покое, вне работы, мышцы никогда полностью не расслабляются, а сохраняют некоторое напряжение, называемое тонусом. Внешне это выражается в упругости мышц. Тонус скелетных мышц связан с поступлением к мышце отдельных следующих друг за другом с большим интервалом нервных импульсов, возбуждающих попеременно различные мышечные волокна. Эти импульсы возникают в мотонейронах спинного мозга, активность которых в свою очередь поддерживается импульсами, исходящими как из вышележащих центров, так и с периферии от рецепторов растяжения («мышечных веретен»), находящихся в самих мышцах.
У человека тонус мышц в известных пределах может регулироваться произвольно: по желанию человек может почти полностью расслабить мышцы или же несколько напрячь их, не совершая, однако, при этом движения.
Работа и сила мышц. Величина сокращения (степень укорочения) мышцы при данной силе раздражения зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект, при сильном же растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения падает.
Для измерения силы мышцы определяют тот максимальный груз, который она в состоянии поднять. Эта сила может быть очень велика: собака, например, мышцами челюсти может поднять груз, превышающий вес ее тела в 8,3 раза. О силе икроножных мышц человека судят по величине груза, положенного ему на плечи, с которым он в состоянии приподняться на носки.
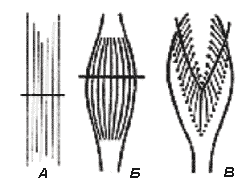
Рис. 6. Типы строения различных мышц (по
А.А. Ухтомскому).
А – мышцы с параллельным ходом волокон; Б –
веретенообразная мышца; В – перистая мышца
Сила мышцы при прочих равных условиях зависит не от ее длины, а от поперечного сечения: чем больше физиологическое поперечное сечение мышцы, т.е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косыми волокнами сумма поперечных сечений волокон может значительно превышать геометрическое поперечное сечение самой мышцы (рис. 6). По этой причине сила мышцы с косыми волокнами значительно больше, чем сила мышцы той же толщины, но с продольными волокнами. Чтобы сравнить силу разных мышц, максимальный груз, который мышца в состоянии поднять, делят на площадь ее поперечного сечения, вычисляя абсолютную мышечную силу. Например, абсолютная сила икроножной мышцы человека равна 5,9 кг/см 2 , сгибателя плеча – 8,1, жевательной мышцы – 10, двуглавой мышцы плеча – 11,4, трехглавой мышцы плеча – 16,8, гладких мышц – 1 кг/см 2 .
Большинство мышц млекопитающих и человека имеет перистое строение. Перистая мышца имеет большое физиологическое сечение, а поэтому обладает большой силой.
Работа мышцы измеряется произведением поднятого груза на величину укорочения мышцы, т.е. выражается в килограммометрах или граммсантиметрах.
По содержанию миоглобина мышечные волокна разделяют на красные, белые и промежуточные. Красные волокна принято считать «медленными», а белые – «быстрыми». Некоторые ученые считают, что при рождении мышцы человека состоят только из «медленных» волокон и часть из них в процессе развития превращается в «быстрые». Другие убеждены, что расположение мышц, особенности их строения и функций предопределены генетически.
Красные волокна работают в основном в аэробном режиме, а белые – в условиях недостатка кислорода, т.е. в них протекают разные метаболические процессы. Красные волокна, как правило, используются для выполнения легкой или умеренной работы, а белые начинают функционировать лишь после значительного возрастания притока возбуждающих импульсов во время очень интенсивной работы. Волокна промежуточного типа сохраняют свойства и красных, и белых волокон и называются еще «быстрыми красными».
Процентное содержание тех или иных волокон предопределяет специализацию атлета. Как правило, обладатели преимущественно красной мускулатуры достигают лучших результатов в видах спорта, требующих повышенной выносливости (плавание, велосипедный спорт, бег на средние и длинные дистанции и т.д.). Те, у кого больше белых мышечных волокон, лучше выполняют силовые упражнения. Последнее объясняется и тем, что белые волокна легче гипертрофируются (увеличиваются в объеме). Однако не все предопределяется только природой. Существуют еще и тренировочные факторы, которым некоторые специалисты отдают предпочтение в формировании структуры мышц.
Как мышцы «привыкают» к физическим нагрузкам. Во время тренировок в мышцах формируются механизмы запасания и использования энергетических субстратов: креатинфосфата, гликогена и жиров (в виде триацилглицеридов). Запасов АТФ и креатинфосфата, или фосфагена, в мышце очень мало. Фосфаты постоянно синтезируются (содержание креатинфосфата в белых мышечных волокнах составляет примерно 30 мкмоль на 1 г сырой массы мышцы), но быстро расходуются при нагрузке, продолжающейся более нескольких секунд. Энергоемкие гликоген и триацилглицериды составляют основную массу резервных источников «топлива» для мышц.
Под влиянием скоростной и силовой тренировки в белых мышечных волокнах повышается активность гликолитических ферментов и образуется большое количество гликогена, который запасается в мышцах в виде гранул, вокруг которых размещаются соответствующие ферменты. Происходят изменения и в буферных системах мышечных клеток: как только внутри клетки при интенсивной работе изменяется рН, это сразу же сказывается на работе ферментов, отвечающих за гликолиз.
Общее количество работы, которое анаэробный гликолиз может обеспечить при интенсивной нагрузке, зависит от резерва гликогена (при распаде каждой его молекулы образуется 6,2 моль АТФ). Само использование запасов гликогена в мышцах запускается гормональными и нервными стимулами. Один из таких гормонов – адреналин – способен значительно активизировать процессы использования гликогена для ресинтеза АТФ.
Под влиянием тренировки анаэробный гликолиз в мышцах можно многократно увеличить. Так, по некоторым данным, у тренированных спринтеров процессы гликолиза в мышцах ног усиливаются в две тысячи раз.
Однако за счет анаэробного гликолиза человек способен выполнять нагрузку только 2–3 минуты. После этого неизбежно запускаются процессы окислительного фосфорилирования.
При длительной работе главными действующими лицами становятся красные и промежуточные мышечные волокна. Энергия для деятельности этих мышц образуется в митохондриях (их в клетках красных мышечных волокон гораздо больше, чем в белых) с помощью окислительных ферментов в присутствии достаточного количества кислорода.
Гликоген, который активно используется при кратковременной работе, является (вместе с жирами) основным эндогенным субстратом и при продолжительной нагрузке. Оба эти вида «топлива», особенно жиры, содержатся в виде запасов в красных и промежуточных волокнах. Жиры несколько уступают гликогену в эффективности выхода энергии на единицу потребляемого кислорода: при их окислении образуется 5,6 моль АТФ.
При окислительном фосфорилировании мышца может получать энергетические субстраты и из центральных депо (гликоген из печени и жир из жировой ткани) и даже использовать энергетические источники, поступающие извне во время работы, например углеводные добавки при марафонском забеге, – природа предусмотрела для длительной работы дополнительные возможности.
Для представителей видов спорта, которые тренируются на выносливость, будет любопытен тот факт, что длительная и интенсивная работа мышц обеспечивается энергией лучше всего при одновременном использовании углеводов и жиров. Казалось бы, здесь некий парадокс. Ведь углеводов хватает только на 20–30 мин интенсивной работы, а жиры могут использоваться гораздо дольше. Однако дело все в том, что использование одних лишь жиров обеспечивает вдвое меньшую скорость выработки энергии, чем одновременное использование жиров и углеводов. А от этого зависит интенсивность выполняемой работы. Таким образом, как считают биохимики, гликоген является наилучшим «топливом» для обеспечения высокоинтенсивной продолжительной работы в аэробных условиях. Ученые нашли даже зависимость длительности работы до полного изнеможения от содержания гликогена в мышцах перед началом нагрузки. Однако, если нагрузка продолжается 2–3 ч, то организм начинает использовать для обеспечения мышечных сокращений и гликоген, и жиры. При переходе на расщепление жиров мощность работы снижается. Вначале используются триацилглицериды, а затем свободные жирные кислоты, которые поступают из крови.
Атлетов, специализирующихся в силовых видах спорта, и культуристов, конечно же, интересует вопрос рабочей гипертрофии мышц.
Систематическая интенсивная работа мышцы приводит к увеличению массы мышечной ткани. Это явление названо рабочей гипертрофией мышцы. В основе гипертрофии лежит увеличение массы протоплазмы мышечных волокон, приводящее к их утолщению. При этом повышается содержание белков и гликогена, а также веществ, доставляющих энергию, необходимую для мышечного сокращения – АТФ и креатинфосфата. Поэтому сила и скорость сокращения гипертрофированной мышцы выше, чем негипертрофированной.
Увеличение массы мышечной ткани у тренированных людей приводит к тому, что мускулатура тела может составлять 50% веса тела (вместо обычных 35–40%).
Гипертрофия развивается, если человек ежедневно на протяжении длительного времени производит мышечную работу, требующую большого напряжения (силовая нагрузка). Мышечная работа, производимая без особых усилий, даже если она продолжается очень долго, к гипертрофии мышцы не приводит.
Гипертрофия мышц очень важна для выполнения кратковременной «взрывной» работы. Возможно, это связано с тем, что запасы креатинфосфата в мышцах человека не увеличиваются больше определенного количества на единицу массы мышц. Таким образом, увеличение объема мышц способствует увеличению общего количества этого энергоемкого субстрата в мышцах и, соответственно, увеличению способности более эффективно выполнять работу максимальной мощности.
Противоположным рабочей гипертрофии явлением служит атрофия мышцы от бездеятельности. Она развивается во всех случаях, когда мышца почему-либо утрачивает способность совершать свою нормальную работу, например, при длительном обездвиживании конечности в гипсовой повязке, при долгом пребывании больного в постели, при перерезке сухожилия, вследствие чего мышца перестает совершать работу против нагрузки, и т.д.
При атрофии диаметр мышечных волокон и содержание в них сократительных белков, гликогена, АТФ и других важных для сократительной деятельности веществ резко падают. При возобновлении нормальной работы мышцы атрофия постепенно исчезает.
Утомление мышц. Временное понижение работоспособности мышц, наступающее в результате работы или тренировки и исчезающее после отдыха, определяется утомлением.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен небольшой груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля. Полученная таким образом кривая называется кривой утомления.
Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения и понижается возбудимость. Но все эти изменения возникают не тотчас же после начала работы мыш-цы – существует некоторый период, в течение которого наблюдаются увеличение амплитуд сокращений и небольшое повышение возбудимости мышцы. При этом мышца становится легко растяжимой. В таких случаях говорят, что мышца «врабатывается», т.е. приспосабливается к работе при заданном ритме и силе раздражения. При дальнейшем длительном раздражении наступает утомление мышечных волокон.
Это может быть обусловлено накоплением в мышце продуктов обмена веществ (в частности, молочной кислоты, образующейся при расщеплении гликогена), оказывающих угнетающее влияние на работоспособность мышечных волокон. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное пространство и оказывают угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия.
Кроме того, на развитие утомления в мышце влияет постепенное истощение в ней энергетических запасов.
Все описанное выше относится к изолированной мышце. Ведь при работе в организме мышца непрерывно снабжается кровью и, следовательно, получает определенное количество питательных веществ (глюкозу, аминокислоты) и освобождается от продуктов обмена, нарушающих нормальную жизнедеятельность мышечных волокон. Главное отличие состоит в том, что в организме возбуждающие импульсы приходят к мышце с нерва. Нервно-мышечное соединение утомляется значительно раньше, чем мышечные волокна, в связи с чем блокирование передачи возбуждения с нерва на мышцу предохраняет последнюю от истощения, вызываемого длительной работой. В целостном организме еще раньше нервно-мышечных соединений утомляются при работе нервные центры.
Восстановление работоспособности утомленных мышц руки человека после длительной работы по подъему груза можно ускорить, если во время отдыха производить работу другой рукой или нижними конечностями.
Продолжение следует
На вопрос Чем обусловлено появление кальция в цитоплазме клеток скелетных мышц? заданный автором роскошествовать
лучший ответ это кальций является фактором, разрешающим сокращение мышц: при повышении концентрации ионов кальция. в миоплазме происходит присоединение Сa к регуляторному белку, в результате чего актин становится способным взаимодействовать с миозином; соединяясь, эти два белка образуют актомиозин, и мышца сокращается. В процессе образования актомиозина происходит расщепление АТФ, химическая энергия которого обеспечивает выполнение механической работы и частично рассеивается в виде тепла. Наибольшая сократительная активность скелетной мышцы наблюдается при концентрации кальция 10-6-10(минус в) -7 моль; при понижении концентрации ионов Ca (менее 10-7 моль) мышечное волокно теряет способность к укорочению и напряжению. Действие Ca на ткани проявляется в изменении их трофики, интенсивности окислительно-восстановительных процессов и в других реакциях, связанных с образованием энергии. Изменение концентрации Ca в омывающей нервную клетку жидкости существенно влияет на проницаемость ее мембраны для ионов калия и особенно для ионов натрия, причем понижение уровня Ca вызывает повышение проницаемости мембраны для ионов натрия и повышение возбудимости нейрона. Повышение концентрации Ca оказывает стабилизирующее влияние на мембрану нервной клетки. Установлена роль Ca в процессах, связанных с синтезом и выделением нервными окончаниями медиаторов, обеспечивающих синаптическую передачу нервного импульса.
Перенос молекул и ионов против электрохимического градиента (активный транспорт) связан со значительными затратами энергии. Часто градиенты достигают больших величин. например, концентрационный градиент водородных ионов на плазматической мембране клеток слизистой оболочки желудка составляет 10в6степени, градиент концентрации ионов кальция на мембране саркоплазматического ретикулума - 10в4 степени, при этом потоки ионов против градиента значительны. В результате затраты энергии на транспортные процессы достигают, например, у человека, более 1/3 всей энергии метаболизма. В плазматических мембранах клеток различных органов обнаружены системы активного транспорта ионов натрия и калия - натриевый насос. Эта система перекачивает натрий из клетки и калий в клетку (антипорт) против их электрохимических градиентов. Перенос ионов осуществляется основным компонентом натриевого насоса - Na+, К+-зависимой АТФ-азой за счет гидролиза АТФ. На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия. Существуют два типа Са2+-АТФ-аз. Одна из них обеспечивает выброс ионов кальция из клетки в межклеточную среду, другая - аккумуляцию кальция из клеточного содержимого во внутриклеточное депо. Обе системы способны создавать значительный градиент иона кальция. К+, Н+-АТФ-аза обнаружена в слизистой оболочке желудка и кишечника. Она способна транспортировать Н+ через мембрану везикул слизистой оболочки при гидролизе АТФ. В микросомах слизистой оболочки желудка лягушки найдена аниончувствительная АТФ-аза, способная при гидролизе АТФ осуществлять антипорт бикарбоната и хлорида.
Если мышцу стимулировать коротким электрическим импульсом, спустя небольшой латентный период происходит ее сокращение. Такое сокращение называется «одиночное сокращение мышцы». Одиночное мышечное сокращение длится около 10-50 мс, причем оно достигает максимальной силы через 5-30 мс.
Каждое отдельное мышечное волокно подчиняется закону «все или ничего», т. е. при силе раздражения выше порогового уровня происходит полное сокращение с максимальной для данного волокна силой, а ступенчатое повышение силы сокращения по мере увеличения силы раздражения невозможно. Поскольку смешанная мышца состоит из множества волокон с различным уровнем чувствительности к возбуждению, сокращение всей мышцы может быть ступенчатым в зависимости от силы раздражения, при этом при сильных раздражениях происходит активация глубжележащих мышечных волокон.
Механизм скольжения филаментов
Укорочение мышцы происходит за счет укорочения образующих ее саркомеров, которые, в свою очередь, укорачиваются за счет скольжения относительно друг друга актиновых и миозиновых филаментов (а не укорочения самих белков). Теория скольжения филаментов была предложена учеными Huxley и Hanson (Huxley, 1974; рис. 1). (В 1954 г. две группы исследователей - X. Хаксли с Дж. Хэнсон и А. Хаксли с Р. Нидергерке - сформулировали теорию, объясняющую мышечное сокращение скольжением нитей. Независимо друг от друга они обнаружили, что длина диска А оставалась постоянной в расслабленном и укороченном саркомере. Это позволило предположить, что есть два набора нитей - актиновые и миозиновые, причем одни входят в промежутки между другими, и при изменении длины саркомера эти нити каким-то образом скользят друг по другу. Сейчас эта гипотеза принята почти всеми.)
Актин и миозин - два сократительных белка, которые способны вступать в химическое взаимодействие, приводящее к изменению их взаимного расположения в мышечной клетке. При этом цепочка миозина прикрепляется к актиновой нити с помощью целого ряда особых «головок», каждая из которых сидит на длинной пружинистой «шее». Когда происходит сцепление между миозиновой головкой и актиновой нитью, конформация комплекса этих двух белков изменяется, миозиновые цепочки продвигаются между актиновыми нитями и мышца в целом укорачивается (сокращается). Однако, чтобы химическая связь между головкой миозина и активной нитью образовалась, необходимо подготовить этот процесс, поскольку в спокойном (расслабленном) состоянии мышцы активные зоны белка актина заняты другим белком - тропохмиозином, который не позволяет актину вступить во взаимодействие с миозином. Именно для того, чтобы убрать тропомиозиновый «чехол» с актиновой нити, требуется быстрое выливание ионов кальция из цистерн саркоплазматического ретикулума, что происходит в результате прохождения через мембрану мышечной клетки потенциала действия. Кальций изменяет конформацию молекулы тропомиозина, в результате чего активные зоны молекулы актина открываются для присоединения головок миозина. Само это присоединение осуществляется с помощью так называемых водородных мостиков, которые очень прочно связывают две белковые молекулы - актин и миозин - и способны в таком связанном виде находиться очень долго.
Для отсоединения миозиновой головки от актина необходимо затратить энергию аденозинтрифосфа-та (АТФ), при этом миозин выступает в роли АТФазы (фермента, расщепляющего АТФ). Расщепление АТФ на аденозиндифосфат (АДФ) и неорганический фосфат (Ф) высвобождает энергию, разрушает связь между актином и миозином и возвращает головку миозина в исходное положение. В дальнейшем между актином и миозином могут снова образовываться поперечные связи.
При отсутствии АТФ актин-миозиновые связи не разрушаются. Это и является причиной трупного окоченения (rigor mortis) после смерти, т. к. останавливается выработка АТФ в организме - АТФ предотвращает мышечную ригидность.
Даже при мышечных сокращениях без видимого укорочения (изометрические сокращения, см. выше) активируется цикл формирования поперечных связей, мышца потребляет АТФ и выделяет тепло. Головка миозина многократно присоединяется на одно и то же место связывания актина, и вся система миофиламентов остается неподвижной.
Внимание : Сократительные элементы мышц актин и миозин сами по себе не способны к укорочению. Мышечное укорочение является следствием взаимного скольжения миофиламентов относительно друг друга (механизм скольжения филаментов).
Как же образование поперечных связей (водородных мостиков) переходит в движение? Одиночный саркомер за один цикл укорачивается приблизительно на 5-10 нм, т.е. примерно на 1 % своей общей длины. За счет быстрого повторения цикла поперечных связей возможно укорочение на 0,4 мкм, или 20% своей длины. Поскольку каждая миофибрилла состоит из множества саркомеров и во всех них одновременно (но не синхронно) образуются поперечные связи, суммарно их работа приводит к видимому укорочению всей мышцы. Передача силы этого укорочения происходит через Z-линии миофибрилл, а также концы сухожилий, прикрепленных к костям, в результате чего и возникает движение в суставах, через которые мышцы реализуют перемещение в пространстве частей тела или продвижение всего тела.
Связь между длиной саркомера и силой мышечных сокращений
Наибольшую силу сокращений мышечные волокна развивают при длине 2-2,2 мкм. При сильном растяжении или укорочении саркомеров сила сокращений снижается (рис. 2). Эту зависимость можно объяснить механизмом скольжения филаментов: при указанной длине саркомеров наложение миозиновых и актиновых волокон оптимально; при большем укорочении миофиламенты перекрываются слишком сильно, а при растяжении наложение миофиламентов недостаточно для развития достаточной силы сокращений.
Скорость укорочения мышечных волокон
Скорость укорочения мышцы зависит от нагрузки на эту мышцу (закон Хилла, рис. 3). Она максимальна без нагрузки, а при максимальной нагрузке практически равна нулю, что соответствует изометрическому сокращению, при котором мышца развивает силу, не изменяя своей длины.
Влияние растяжения на силу сокращений: кривая растяжения в покое
Важным фактором, влияющим на силу сокращений, является величина растяжения мышцы. Тяга за конец мышцы и натяжение мышечных волокон называются пассивным растяжением. Мышца обладает эластическими свойствами, однако в отличие от стальной пружины зависимость напряжения от растяжения не линейна, а образует дугообразную кривую. С увеличением растяжения повышается и напряжение мышцы, но до определенного максимума. Кривая, описывающая эти взаимоотношения, называется кривой растяжения в покое .
Данный физиологический механизм объясняется эластическими элементами мышцы - эластичностью сарколеммы и соединительной ткани, располагающимися параллельно сократительным мышечным волокнам.
Также при растяжении изменяется и наложение друг на друга миофиламентов, однако это не оказывает влияния на кривую растяжения, т. к. в покое не образуются поперечные связи между актином и миозином. Предварительное растяжение (пассивное растяжение) суммируется с силой изометрических сокращений (активная сила сокращений).
Читайте также
рис. 1. Схема образования поперечных связей - молекулярной основы сокращения саркомера

рис. 2. Зависимость силы сокращений от длины саркомера

рис.3. Зависимость скорости укорочения от нагрузки

рис. 4. Влияние предварительного растяжения на силу сокращения мышцы. Предварительное растяжение повышает напряжение мышцы. Результирующая кривая, описывающая взаимоотношения длины мышцы и силы ее сокращения при воздействии активного и пассивного растяжения, демонстрирует более высокое изометрическое напряжение, чем в покое
Одиночное сокращение, суммация, тетанус.
При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
1.Латентный период. Это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мсек. Во время латентного периода генерируется и распространяется ПД, происходит высвобождения кальция из СР, взаимодействие актина с миозином и т.д.
2.Период укорочения. В зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек.,
3.Период расслабления. Его длительность несколько больше, чем укорочения. Рис.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например так могут сокращаться быстрые глазодвигательные мышцы. Чаще одиночные сокращения суммируются.
Суммация это сложение 2-х последовательных сокращений при нанесении на нее 2-х пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефрактерного периода. Различают 2 вида суммации: полную и неполную суммацию. Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда он уже начала расслабляться. Полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е.
Мышечное сокращение
в конце периода укорочения.(рис 1,2). Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить. Например нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности.
Тетанус- это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий. Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация (рис). Гладкий тетанус возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения. Т.е. имеет место полная суммация отдельных сокращений и (рис). Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например тремор рук при алкогольной интоксикации и болезни Паркинсона.
− В зависимости от условий , в которых происходит мышечное сокращение, различают два его основных типа - изотоническое и изометрическое . Сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным, называется изотоническим . Изометрическим является такое сокращение, при котором мышца укоротиться не может, если оба ее конца закреплены неподвижно.
Механизм мышечного сокращения
В этом случае по мере развития сократительного процесса напряжение возрастает, а длина мышечных волокон остается неизменной.
В натуральных двигательных актах сокращения мышц смешанные: даже поднимая постоянный груз, мышца не только укорачивается, но и изменяет свое напряжение вследствие реальной нагрузки. Такое сокращение называется ауксотоническим.
− В зависимости от частоты стимуляции выделяют одиночные и тетанические сокращения.
Одиночное сокращение (напряжение) возникает при действии на мышцу одиночного электрического или нервного импульса. Волна возбуждения возникает в месте приложения электродов для прямого раздражения мышцы или в области нервно-мышечного соединения и отсюда распространяется вдоль всего мышечного волокна. В изотоническом режиме одиночное сокращение икроножной мышцы лягушки начинается через короткий скрытый (латентный) период - до 0,01 с, далее следуют фаза подъема (фаза укорочения) - 0,05 с и фаза спада (фаза расслабления) - 0,05-0,06 с. Обычно мышца укорачивается на 5-10 % от исходной длины. Как известно, продолжительность волны возбуждения (ПД) мышечных волокон варьирует, составляя величину порядка 1-10 мс (с учетом замедления фазы реполяризации в ее конце). Таким образом, длительность одиночного сокращения мышечного волокна, наступающего вслед за его возбуждением, во много раз превышает продолжительность ПД.
Мышечное волокно реагирует на раздражение по правилу «все или ничего», т.е. отвечает на все надпороговые раздражения стандартным ПД и стандартным одиночным сокращением. Однако сокращение целой мышцы при ее прямом раздражении находится в большой зависимости от силы стимуляции. Это связано с различной возбудимостью мышечных волокон и разным расстоянием их от раздражающих электродов, что ведет к неодинаковому количеству активированных мышечных волокон.
При пороговой силе стимула сокращение мышцы едва заметно, потому что в реакцию вовлекается лишь небольшое количество волокон. При увеличении силы раздражения число возбужденных волокон растет, пока все волокна не окажутся сокращенными, и тогда достигается максимальное сокращение мышцы. Дальнейшее усиление стимулов прироста амплитуды сокращения не вызывает.
В естественных условиях мышечные волокна работают в режиме одиночных сокращений только при относительно низкой частоте импульсации мотонейронов, когда интервалы между последовательными ПД мотонейронов превышают длительность одиночного сокращения иннервируемых ими мышечных волокон. Еще до прихода следующего импульса от мотонейронов мышечные волокна успевают полностью расслабиться. Новое сокращение возникает после полного расслабления мышечных волокон. Такой режим работы обусловливает незначительную утомляемость мышечных волокон. При этом ими развивается относительно небольшое напряжение.
Тетаническое сокращение - это длительное слитное сокращение скелетных мышц. В его основе лежит явление суммации одиночных мышечных сокращений. При нанесении на мышечное волокно или целую мышцу двух быстро следующих друг за другом раздражении возникающее сокращение будет иметь большую амплитуду. Сократительные эффекты, вызванные первым и вторым раздражениями, как бы складываются, происходит суммация, или суперпозиция, сокращений, поскольку нити актина и миозина дополнительно скользят друг относительно друга. При этом в сокращение могут вовлекаться ранее не сокращавшиеся мышечные волокна, если первый стимул вызвал у них подпороговую деполяризацию, а второй увеличивает ее до критической величины. При получении суммации в одиночном волокне важно, чтобы второе раздражение наносилось после исчезновения ПД, т.е. после рефрактерного периода. Естественно, что суперпозиция сокращений наблюдается и при стимуляции моторного нерва, когда интервал между раздражениями короче всей длительности сократительного ответа, в результате чего и происходит слияние сокращений.
При сравнительно низких частотах наступает зубчатый тетанус , при большой частоте - гладкий тетанус (рис. 13).
Рис. 13. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва. Суперпозиция волн сокращения и образование разных видов тетануса.
а - одиночное сокращение (Г = 1 Гц); б, в - зубчатый тетанус (Г= 15-20 Гц); г, д - гладкий тетанус и оптимум (Г = 25-60 Гц); е - пессимум - расслабление мышцы во время раздражения (Г= 120 Гц).
Их амплитуда больше величины максимального одиночного сокращения. Напряжение, развиваемое мышечными волокнами при гладком тетанусе, обычно в 2-4 раза больше, чем при одиночном сокращении. Режим тетанического сокращения мышечных волокон в отличие от режима одиночных сокращений быстрее вызывает их утомление и поэтому не может поддерживаться длительное время. Из-за укорочения или полного отсутствия фазы расслабления мышечные волокна не успевают восстановить энергетические ресурсы, израсходованные в фазе укорочения. Сокращение мышечных волокон при тетаническом режиме с энергетической точки зрения происходит «в долг».
До сих пор нет общепризнанной теории, объясняющей, почему напряжение, развиваемое при тетанусе, или суперпозиции сокращений, гораздо больше, чем при одиночном сокращении. Во время кратковременной активации мышцы вначале одиночного сокращения в поперечных мостиках между нитями актина и миозина возникает упругое напряжение. Однако недавно было показано, что такой активации недостаточно для прикрепления всех мостиков. Когда она более длительная, обеспечиваемая ритмической стимуляцией (например, при тетанусе), их прикрепляется больше. Количество поперечных мостиков, связывающих актиновые и миозиновые филаменты (а, следовательно, и развиваемая мышцей сила), согласно теории скользящих нитей, зависит от степени перекрывания толстых и тонких нитей, а значит, и от длины саркомера или мышцы.
Высвобождение Са 2+ при тетанусе . Если стимулы поступают с высокой частотой (не менее 20 Гц), уровень Са 2 + в интервалах между ними остается высоким, потому что кальциевый насос не успевает вернуть все ионы в продольную систему саркоплазматического ретикулума. В таких условиях отдельные сокращения почти полностью сливаются. Это состояние устойчивого сокращения, или тетанус, наблюдается в том случае, когда промежутки между стимулами (или потенциалами действия в клеточной мембране) меньше примерно 1/3 длительности каждого из одиночных сокращений. Следовательно, частота стимуляции, необходимая для их слияния, тем ниже, чем больше их длительность; по этой причине она зависит от температуры. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода, который приблизительно соответствует длительности потенциала действия.
Как выяснилось, амплитуда гладкого тетануса колеблется в широких пределах в зави- \ симости от частоты стимуляции нерва. При некоторой оптимальной (достаточно высокой) частоте стимуляции амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус получил название оптимума . При дальнейшем повышении частоты стимуляции нерва развивается блок проведения возбуждения в нервно-мышечных синапсах, приводящий к расслаблению мышцы в ходе стимуляции нерва - пессимум Введенского. Частота стимуляции нерва, при которой наблюдается пессимум, получила название пессимальной (см. рис. 6.4).
В эксперименте легко обнаруживается, что уменьшенная в ходе пессимальной ритмической стимуляции нерва амплитуда мышечного сокращения моментально возрастает при возвращении частоты раздражения от пессимальной к оптимальной. В этом наблюдении - хорошее доказательство того, что пессимальное расслабление мышцы не является следствием утомления, истощения энергоемких соединений, а является следствием особых соотношений, складывающихся на уровне пост- и пресинаптических структур нервно-мышечного синапса. Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (около 200 имп/с).
Контрактура . Контрактурой называется состояние обратимого местного устойчивого сокращения. Оно отличается от тетануса отсутствием распространяющегося потенциала действия. При этом может наблюдаться длительная локальная деполяризация мышечной мембраны, например при калиевой контрактуре, или же мембранный потенциал, близкий к уровню покоя, в частности при кофеиновой контрактуре. Кофеин при нефизиологически высоких (миллимолярных) концентрациях проникает в мышечные волокна и, не вызывая возбуждения мембраны, способствует высвобождению Са 2+ из саркоплазматического ретикулума; в результате развивается контрактура.
При калиевой контрактуре степень стойкой деполяризации и сократительного напряжения волокна зависит от концентрации К + в наружном растворе.
⇐ Предыдущая32333435363738394041Следующая ⇒
Дата публикования: 2015-07-22; Прочитано: 5124 | Нарушение авторского права страницы
Studopedia.org — Студопедия.Орг — 2014-2018 год.(0.001 с)…
Мышечное сокращение
Конспект лекции | Резюме лекции | Интерактивный тест | Скачать конспект
» Структурная организация скелетной мышцы
» Молекулярные механизмы сокращения скелетной мышцы
» Сопряжение возбуждения и сокращения в скелетной мышце
» Расслабление скелетной мышцы
»
» Работа скелетной мышцы
» Структурная организация и сокращение гладких мышц
» Физиологические свойства мышц
Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движения глазных яблок, глотание, дыхание и т.
п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл, которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты .
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белка актина . В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку и хвост . Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик .
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки ), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++ .
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ) . ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона.
Физиология скелетных мышц
Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей , мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга.
Механизм скольжения нитей включает несколько последовательных событий.
Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.
В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения .
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
Латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
Фаза укорочения (около 50 мс);
Фаза расслабления (около 50 мс).
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
Числом ДЕ, участвующих в сокращении;
Частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов.
Структурная организация и сокращение гладких мышц
Структурная организация. Гладкая мышца состоит из одиночных клеток веретенообразной формы (миоцитов ), которые располагаются в мышце более или менее хаотично. Сократительные филламенты расположены нерегулярно, вследствие чего отсутствует поперечная исчерченность мышцы.
Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре.
Механизм сопряжения возбуждения и сокращения.
При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам.
Сокращение и расслабление гладких мышц. Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам.
Физиологические свойства мышц
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость . Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1. Физиологические свойства и особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1. Сравнительная характеристика скелетных и гладких мышц
|
Свойство |
Скелетные мышцы |
Гладкие мышцы |
|
Скорость деполяризации |
медленная |
|
|
Период рефрактерности |
короткий |
длительный |
|
Характер сокращения |
быстрые фазические |
медленные тонические |
|
Энергозатраты |
||
|
Пластичность |
||
|
Автоматия |
||
|
Проводимость |
||
|
Иннервация |
мотонейронами соматической НС |
постганглионарными нейронами вегетативной НС |
|
Осуществляемые движения |
произвольные |
непроизвольные |
|
Чувствительность к химическим веществам |
||
|
Способность к делению и дифференцировке |
Пластичность гладких мышц проявляется в том, что они могут сохранять постоянный тонус как в укороченном, так и в растянутом состоянии.
Проводимость гладкой мышечной ткани проявляется в том, что возбуждение распространяется от одного миоцита к другому через специализированные электропроводящие контакты (нексусы).
Свойство автоматии гладкой мускулатуры проявляется в том, что она может сокращаться без участия нервной системы, за счет того, что некоторые миоциты способны самопроизвольно генерировать ритмически повторяющиеся потенциалы действия.
Двигательная система человека. Концентрическое сокращение

Цикл удлинения-сокращения. Общей схемой возбуждения мышц, в частности, во время выполнения задач, требующих высокой напряженности, является использование концентрической последовательности, в которой действующая мышца сначала удлиняется, а затем сокращается.
Преимущество этой стратегии заключается в том, что мышца может выполнить большую положительную работу, если она активно растягивается, прежде чем сократится. В результате этого цикла удлинения-сокращения во время концентрического сокращения выполняется большее количество работы по сравнению с тем, если бы мышца сама по себе выполняла концентрическое сокращение.
Экспериментальное доказательство этого вывода основывается на работе, выполняемой отдельной мышцей. Эксперимент состоял из двух частей: сначала мышца растягивалась, а затем стимулировалась, прежде чем сократиться и выполнить положительную работу; после этого мышца сначала стимулировалась, а затем растягивалась, прежде чем выполнить положительную работу.
Результаты каждой части эксперимента показаны в виде графиков длина-время, усилие-время и усилие-длина. Критическое сравнение выполняется на графиках усилие-длина. Фаза с показывает изменение усилия и длины при выполнении мышцей работы. Поскольку работа определяется как произведение усилия и смещения, площадь ниже кривой усилие-длина во время фазы с представляет работу, выполняемую в течение каждой части эксперимента.
Безусловно, площадь ниже этой кривой больше для второй части эксперимента, которая состояла из растягивания (удлинения) действующей мышцы; это соответствует циклу удлинения-сокращения. Соотношение между работой и энергией свидетельствует, что увеличение работы, выполняемой мышцей, требует повышения расхода энергии.
Откуда могла бы поступить эта дополнительная энергия?
Механизмы сокращения скелетных мышц
Типичное двухэлементное логическое обоснование таково. Во-первых, эксцентрическое сокращение нагружает последовательный упругий элемент в результате его растяжения, что может быть представлено в качестве передачи энергии от нагрузки последовательному упругому элементу; это представляет собой накопление упругой энергии.
Например, если удерживать упругую ленту одним концом в каждой руке и затем растянуть, действие мышц кистей рук, участвующих в растягивании ленты, накапливается в ленте в виде упругой энергии. Во-вторых, после освобождения молекулярная структура упругой ленты использует эту упругую энергию для возвращения к своей первоначальной форме.
Аналогичным образом, поскольку отношение мышечной силы к усилию нагрузки изменяется и мышца подвергается концентрическому сокращению, упругая энергия, накопленная в последовательном упругом элементе, может быть восстановлена и использована для содействия укорачивающему сокращению (положительная работа). При возбуждении мышцы в результате множества процессов обмена веществ в качестве существенного элемента химической энергии образуется АТФ.
В процессе как образования так и использования некоторая часть энергии расходуется в виде тепла. Согласно приведенному выше уравнению, если используемая химическая энергия и выделяющееся тепло остаются постоянными (допустим, для данного случая EPiS = 0), количество выполненной работы останется таким же.
Тем не менее выполняемая работа увеличивается при осуществлении эксцентрически концентрической последовательности (удлинение-сокращение). Объясняется это тем, что изменяется либо Е. Однако, согласно объяснению, основывающемуся на явлении накопления и использования упругой энергии, для осуществления работы предоставляется дополнительная энергия наряду с той, которая предоставляется химическими средствами.
На эту способность использовать накапливаемую упругую энергию оказывают влияние три переменные: время, величина удлинения и скорость удлинения. Вероятно, потеря энергии обусловливается разъединением и восстановлением во время этой задержки поперечных межклеточных мостиков, вследствие чего после восстановления миофиламенты испытывают меньшее натяжение.
Аналогично если удлиняющее сокращение слишком большое, после удлинения сохраняется меньше поперечных межклеточных мостиков и, следовательно, накапливается меньше упругой энергии. Однако при условии, что поперечные межклеточные мостики сохраняются, чем выше скорость удлинения, тем больше накапливается упругой энергии (например, Rack, Westbury, 1974).
Несмотря на широко распространенное использование явления накопления и использования упругой энергии для учета увеличения положительной работы, связанной с эксцентрическо-концентрическими сокращениями, увеличение положительной работы обусловливается, вероятно, также значительным возрастанием количества предоставляемой химической энергии.
Это увеличение предоставляемой химической энергии называется пред нагрузочным эффектом. Заметим, например, что в начале фазы с на графике усилие-длина, усилие больше во время эксцентрически концентрического состояния, чем во время изометрическо-концентрического состояния; это соответствует крайне правому пику на графиках усилие-длина.
Безусловно, в начале концентрической фазы усилие в эксцентрически концентрическом состоянии больше. Относительные вклады упругой энергии и пред нагрузочных эффектов можно оценить в результате рассмотрения высоты, которую субъекты могут преодолеть, используя два типа прыжков в высоту (Komi, Bosco, 1978).
Прыжок согнув ноги начинается из положения на корточках (угол коленного сустава около 2 рад) и просто включает разгибание коленных и голеностопных суставов; руки удерживаются вытянутыми над головой для минимизации их вклада в прыжок. Прыжок движением в противоположном направлении начинается из вертикального положения тела и включает в течение одного непрерывного движения приседание до угла коленного сустава около 2 рад и последующее разгибание коленных и голеностопных суставов, как при выполнении прыжка согнув ноги.
Основное различие между двумя этими способами заключается в методике использования сильных экстензоров коленных суставов, выполняющих около 50 % работы во время максимального дружка в высоту (Hublay, Wells, 1983); а именно, прыжок согнув ноги включает только изометрическое концентрическое сокращение экстензоров коленных суставов, в то время как прыжок движением в противоположном направлении требует эксцентрически концентрической последовательности.
zdoroviezubwБлог
Тесты - Следующий тест - На первую страницу
Ответы и объяснения
|
Нервно-мышечная передача возбуждения . Выше мы уже показывали, что проведение возбуждения в нервных и мышечных волокнах осуществляется с помощью электрических импульсов, распространяющихся по поверхностной мембране. Передача же возбуждения с нерва на мышцу основана на другом механизме. Она осуществляется в результате выделения нервными окончаниями высокоактивных химических соединений - медиаторов нервного импульса. В синапсах скелетных мышц таким медиатором является ацетилхолин (АХ).
В нервно-мышечном синапсе выделяют три основных структурных элемента - пресинаптическая мембрана на нерве, постсинаптическая мембрана на мышце, между ними - синаптическая щель . Форма синапса может быть разнообразной. В состоянии покоя АХ содержится в так называемых синаптических пузырьках внутри концевой пластинки нервного волокна. От синаптической щели цитоплазма волокна с плавающими в ней синаптическими пузырьками отделена пресинаптической мембраной. При деполяризации пресинаптической мембраны меняется ее заряд и проницаемость, пузырьки подходят близко к мембране и изливаются в синаптическую щель, ширина которой достигает 200-1000 ангстрем. Медиатор начинает диффундировать через щель к постсинаптической мембране.
Постсинаптическая мембрана не электрогенна, но обладает высокой чувствительностью к медиатору за счет наличия в ней так называемых холинорецепторов - биохимических групп, способных избирательно реагировать с АХ. Последний достигает постсинаптической мембраны через 0,2-0,5 мсек. (так называемая "синаптическая задержка" ) и, взаимодействуя с холинорецепторами, вызывает изменение проницаемости мембраны для Na, что приводит к деполяризации постсинаптической мембраны и генерации на ней волну деполяризации, которая носит название возбуждающего постсинаптического потенциала , (ВПСП ), величина которого превышает Ек соседних, электрогенных участков мембраны мышечного волокна. В результате в них возникает ПД (потенциал действия), который распространяется по всей поверхности мышечного волокна, вызывая затем его сокращение, инициируя процесс т.н. электромеханического сопряжения (Каплинг). Медиатор в синаптической щели и на постсинаптической мембране работает очень короткое время, так как разрушается ферментом холинэстеразой, которая готовит синапс к восприятию новой порции медиатора. Показано также, что часть не прореагировавшего АХ может возвращаться в нервное волокно.
При очень частых ритмах раздражения постсинаптические потенциалы могут суммироваться, так как холинэстераза не успевает полностью расщепить выделяющийся в нервных окончаниях АХ. В результате такой суммации постсинаптическая мембрана все более и более деполяризуется. При этом соседние электрогенные участки мышечного волокна приходят в состояние угнетения, сходное с тем, которое развивается при продолжительном действии катода постоянного тока (катодическая депрессия Вериго).
Функции и свойства поперечно-полосатых мышц.
Поперечно-полосатые мышцы являются активной частью опорно-двигательного аппарата. В результате сократительной деятельности этих мышц происходит перемещение тела в пространстве, перемещение частей тела относительно друг друга, поддержание позы. Кроме того, при мышечной работе вырабатывается тепло.
Каждое мышечное волокно обладает следующими свойствами: возбудимостью , т.е. способностью отвечать на действие раздражителя генерацией ПД, проводимостью - способностью проводить возбуждение вдоль всего волокна в обе стороны от точки раздражения, и сократимостью , т.е. способностью сокращаться или изменять свое напряжение при возбуждении. Возбудимость и проводимость являются функциями поверхностной клеточной мембраны - сарколеммы, а сократимость - функцией миофибрилл, расположенных в саркоплазме.
Методы исследования . В естественных условиях возбуждение и сокращение мышц вызывается нервными импульсами. Для того же, чтобы возбудить мышцу в эксперименте или при клиническом исследовании, ее подвергают искусственному раздражению электрическим током. Непосредственное раздражение самой мышцы называется прямым, а раздражение нерва - непрямым раздражением. Ввиду того, что возбудимость мышечной ткани меньше, чем нервной, приложение электродов непосредственно к мышце еще не обеспечивает прямого раздражения - ток, распространяясь по мышечной ткани, действует в первую очередь на находящиеся в ней окончания двигательных нервов. Чистое прямое раздражение получается лишь при внутриклеточном раздражении или после отравления нервных окончаний кураре. Регистрация мышечного сокращения производится с помощью механических приспособлений - миографов, или специальными датчиками. При изучении мышц используются и электронная микроскопия, регистрация биопотенциалов при внутриклеточном отведении и другие тонкие методики, позволяющие исследовать свойства мышц как в эксперименте, так и в клинике.
Механизмы мышечного сокращения .
Структура миофибрилл и ее изменения при сокращении . Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечно-полосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни из этих участков анизотропны, т.е. обладают двойным лучепреломлением. В обычном свете они выглядят темными, а в поляризованном - прозрачными в продольном и непрозрачными в поперечном направлении. Другие участки изотропны, и выглядят прозрачными при обыкновенном свете. Анизотропные участки обозначаются буквой А , изотропные - I. В середине диска А проходит светлая полоска Н , а посередине диска I проходит темная полоска Z , представляющая собой тонкую поперечную мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию такой опорной структуры параллельно расположенные однозначные диски отдельных миофибрилл внутри одного волокна во время сокращения не смещаются по отношению друг к другу.
Установлено, что каждая из миофибрилл имеет диаметр около 1 мк и состоит в среднем из 2500 протофибрилл, представляющих собой удлиненные полимеризованные молекулы белком миозина и актина. Миозиновые нити (протофибриллы) вдвое толще актиновых. Их диаметр составляет примерно 100 ангстрем. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями. В таком участке каждая толстая нить окружена 6 тонкими. Благодаря этому диски I состоят только из актиновых нитей, а диски А еще и из нитей миозина. Светлая полоска Н представляет собой зону, свободную в период покоя от актиновых нитей. Мембрана Z, проходя через середину диска I, скрепляет между собой нити актина.
Важным компонентом ультрамикроскопической структуры миофибрилл являются также многочисленные поперечные мостики на миозине. В свою очередь на нитях актина имеются так называемые активные центры, в покое прикрытые, как чехлом, специальными белками - тропонином и тропомиозином. В основе сокращения лежит процесс скольжения нитей актина относительно миозиновых нитей. Такое скольжение вызывается работой т.н. "химического зубчатого колеса", т.е. периодически протекающих циклов изменений состояния поперечных мостиков и их взаимодействия с активными центрами на актине. В этих процессах важную роль играют АТФ и ионы Са+.
При сокращении мышечного волокна нити актина и миозина не укорачиваются, а начинают скользить друг по другу: актиновые нити вдвигаются между миозиновыми, в результате чего длина дисков I укорачивается, а диски А сохраняют свой размер, сближаясь друг с другом. Полоска Н почти исчезает, т.к. концы актина соприкасаются и даже заходят друг за друга.
Роль ПД в возникновении мышечного сокращения (процесс электромеханического сопряжения). В скелетной мышце в естественных условиях инициатором мышечного сокращения является потенциал действия, распространяющийся при возбуждении вдоль поверхностной мембраны мышечного волокна.
Если кончик микроэлектрода приложить к поверхности мышечного волокна в области мембраны Z, то при нанесении очень слабого электрического стимула, вызывающего деполяризацию, диски I по обе стороны от места раздражения начнут укорачиваться. при этом возбуждение распространяется вглубь волокна, вдоль мембраны Z. Раздражение других участков мембраны такого эффекта не вызывает. Из этого следует, что деполяризация поверхностной мембраны в области диска I при распространении ПД является пусковым механизмом сократительного процесса.
Дальнейшие исследования показали, что важным промежуточным звеном между деполяризацией мембраны и началом мышечного сокращения является проникновение в межфибриллярное пространство свободных ионов СА++. В состоянии покоя основная часть Са++ в мышечном волокне хранится в саркоплазматическом ретикулюме.
В механизме мышечного сокращения особую роль играет та часть ретикулюма, которая локализована в области мембраны Z. Электронно-микроскопически здесь обнаруживается т.н. триада (Т-система) , каждая из которых состоит из центрально расположенной в области мембраны Z тонкой поперечной трубочки, идущей поперек волокна, и двух боковых цистерн саркоплазматического ретикулюма, в которых заключен связанный Са++. ПД, распространяющийся вдоль поверхностной мембраны, проводится вглубь волокна по поперечным трубочкам триад. Затем возбуждение передается на цистерны, деполяризует их мембрану и она становится проницаема для СА++.
Экспериментально установлено, что существует некоторая критическая концентрация свободных ионов Са++, при которой начинается сокращение миофибрилл. Она равна 0,2-1,5*10 6 ионов на волокно. Увеличение концентрации Са++ до 5*10 6 вызывает уже максимальное сокращение.
Начало мышечного сокращения приурочено к первой трети восходящего колена ПД, когда его величина достигает примерно 50 мв. Полагают, что именно при этой величине деполяризации концентрация Са++ становится пороговой для начала взаимодействия актина и миозина.
Процесс освобождения Са++ прекращается после окончания пика ПД. Тем не менее сокращение продолжает еще нарастать до тех пор, пока не вступает в действие механизм, обеспечивающий возвращение Са++ в цистерны ретикулюма. Такой механизм назван "кальциевым насосом". Для осуществления его работы используется энергия, получаемая при расщеплении АТФ.
В межфибриллярном пространстве Са++ взаимодействует с белками, закрывающими активные центры актиновых нитей - тропонином и тропомиозином, обеспечивая возможность для осуществления реакции поперечных мостиков миозина и нитей актина.
Таким образом, последовательность событий, ведущих к сокращению, а затем к расслаблению мышечного волокна, рисуется в настоящее время так:
Раздражение -- возникновение ПД -- проведение его вдоль клеточной мембраны и вглубь волокна по трубочкам Т-систем -деполяризация мембраны саркоплазматического ретикулюма -- освобождение Са++ из триад и диффузия его к миофибриллам -- взаимодействие Са++ с тропонином и выделение энергии АТФ -- взаимодействие (скольжение) актиновых и миозиновых нитей -- сокращение мышцы -- понижение концентрации Са++ в межфибриллярном пространстве из-за работы Са-насоса -- расслабление мышцы .
Роль АТФ в механизме мышечного сокращения . В процессе взаимодействия актиновых и миозиновых нитей в присутствии ионов Са++ важную роль играет богатое энергией соединение - АТФ. Миозин обладает свойствами фермента АТФ-азы. При расщеплении АТФ освобождается около 10 000 кал. на 1 моль. Под влиянием АТФ изменяются и механические свойства миозиновых нитей - резко увеличивается их растяжимость. Полагают, что расщепление АТФ является источником энергии, необходимой для скольжения нитей. Ионы Са++ повышают АТФ-азную активность миозина. Кроме того, энергия АТФ используется для работы кальциевого насоса в ретикулюме. В соответствии с этим ферменты, расщепляющие АТФ, локализуются в этих мембранах, а не только в миозине.
Ресинтез АТФ, непрерывно расщепляющейся в процессе работы мышц, осуществляется двумя основными путями. Первый состоит в ферментативном переносе фосфатной группы от креатинфосфата (КФ) на АДФ. КФ содержится в мышце в значительно больших количествах, чем АТФ, и обеспечивает ее ресинтез в течение тысячных долей секунды. Однако при длительной работе мышцы запасы КФ истощаются, поэтому важен второй путь - медленный ресинтез АТФ, связанный с гликолизом и окислительными процессами. Окисление молочной и пировиноградной кислот, образующихся в мышце во время ее сокращения, сопровождается фосфорилированием АДФ и креатина, т.е. ресинтезом КФ и АТФ.
Нарушение ресинтеза АТФ ядами, подавляющими гликолиз и окислительные процессы, ведет к полному исчезновению АТФ и КФ, вследствие чего кальциевый насос перестает работать. Концентрация Са++ в области миофибрилл сильно возрастает и мышца приходит в состояние длительного необратимого укорочения - т.н. контрактуры.
Теплообразование при сократительном процессе . По своему происхождению и времени развития теплообразование это делится на две фазы. Первая во много раз короче второй и носит название начального теплообразования. Она начинается с момента возбуждения мышцы и продолжается в течение всего сокращения, включая и фазу расслабления. Вторая фаза теплообразования происходит в течение нескольких минут после расслабления, и носит название запаздывающего, или восстановительного теплообразования. В свою очередь начальное теплообразование может быть разделено на несколько частей - тепло активации, тепло укорочения, тепло расслабления. Тепло, образующееся в мышцах, поддерживает температуру тканей на уровне, обеспечивающем активное протекание физических и химических процессов в организме.
Виды сокращений . В зависимости от условий, в которых происходит сокраще-
ние, различают два его типа - изотоническое и изометрическое . Изотоническим называется такое сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается прежним. Примером является укорочение без нагрузки. Изометрическим называется такое сокращение, при котором мышца укорачиваться не может (когда ее концы неподвижно закреплены). В этом случае длина мышечных волокон остается неизменной, но напряжение их растет (подъем непосильного груза).
Естественные сокращения мышц в организме никогда не бывают чисто изотоническими или изометрическими.
Одиночное сокращение . Раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом вызывает одиночное сокращение мышцы. В нем различают две основные фазы: фазу сокращения и фазу расслабления. Сокращение мышечного волокна начинается уже во время восходящей ветви ПД. Длительность сокращения в каждой точке мышечного волокна в десятки раз превышает продолжительность ПД. Поэтому наступает момент, когда ПД прошел вдоль всего волокна и закончился, волна же сокращения охватила все волокно и оно продолжает быть укороченным. Это соответствует моменту максимального укорочения или напряжения мышечного волокна.
Сокращение каждого отдельного мышечного волокна при одиночных сокращениях подчиняется закону "все или ничего ". Это означает, что сокращение, возникающее как при пороговом, так и при сверхпороговом раздражении, имеет максимальную амплитуду. Величина же одиночного сокращения всей мышцы зависит от силы раздражения. При пороговом раздражении сокращение ее едва заметно, с увеличением же силы раздражения оно нарастает, пока не достигнет известной высоты, после чего уже остается неизменной (максимальное сокращение). Это объясняется тем, что возбудимость отдельных мышечных волокон неодинакова, и поэтому только часть их возбуждается при слабом раздражении. При максимальном сокращении они возбуждены все. Скорость проведения волны сокращения мышцы совпадает со скоростью распространения ПД. В двуглавой мышце плеча она равна 3,5-5,0 м/сек.
Суммация сокращений и тетанус . Если в эксперименте на отдельное мышечное волокно или на всю мышцу действуют два быстро следующих друг за другом сильных одиночных раздражения, то возникающее сокращение будет иметь большую амплитуду, чем максимальное одиночное сокращение. Сократительные эффекты, вызванные первым и вторым раздражением, как бы складываются. Это явление носит название суммации сокращений. Для возникновения суммации необходимо, чтобы интервал между раздражениями имел определенную длительность - он должен быть длиннее рефрактерного периода, но короче всей длительности одиночного сокращения, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться. При этом возможны два случая. Если второе раздражение поступает, когда мышца уже начала расслабляться, на миографической кривой вершина второго сокращения будет отделяться от первого западением. Если же второе раздражение действует, когда первое сокращение еще не дошло до своей вершины, то второе сокращение как бы сливается с первым, образуя вместе с ним единую суммированную вершину. Как при полной, так и при неполной суммации ПД не суммируются. Такое суммированное сокращение в ответ на ритмические раздражения называются тетанусом. В зависимости от частоты раздражения он бывает зубчатый и гладкий.
Причина суммации сокращений при тетанусе кроется в накоплении ионов Са++ в межфибриллярном пространстве до концентрации 5*10 6 мМ/л. После достижения этой величины дальнейшее накопление Са++ не приводит к увеличению амплитуды тетануса.
После прекращения тетанического раздражения волокна вначале расслабляются не полностью, и их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется посттетанической, или остаточной контрактурой. Она связана с тем. что требуется больше времени для удаления из межфибриллярного пространства всего Са++, попавшего туда при ритмических стимулах и не успевшего полностью удалиться в цистерны саркоплазматического ретикулюма работой Са-насосов.
Если после достижения гладкого тетануса еще больше увеличивать частоту раздражения, то мышца при какой-то частоте начинает вдруг расслабляться. Это явление называется пессимумом . Он наступает тогда, когда каждый следующий импульс попадает в рефрактерность от предыдущего.
Моторные единицы . Мы рассмотрели общую схему явлений, лежащих в основе тетанического сокращения. Для того, чтобы более подробно познакомиться с тем, как этот процесс совершается в условиях естественной деятельности организма, необходимо остановиться на некоторых особенностях иннервации скелетной мышцы двигательным нервом.
Каждое моторное нервное волокно, являющееся отростком двигательной клетки передних рогов спинного мозга (альфа-мотонейрона), в мышце ветвиться и иннервирует целую группу мышечных волокон. Такая группа называется моторной единицей мышцы. Количество мышечных волокон, входящих в состав моторной единицы, вариирует в широких пределах, но их свойства одинаковы (возбудимость, проводимость и др.). Вследствие того, что скорость распространения возбуждения в нервных волокнах, иннервирующих скелетные мышцы, очень велика, мышечные волокна, составляющие моторную единицу, приходят в состояние возбуждения практически одновременно. Электрическая активность моторной единицы имеет вид частокола, в котором каждому пику соответствует суммарный потенциал действия многих одновременно возбужденных мышечных волокон.
Следует сказать, что возбудимость различных скелетных мышечных волокон и состоящих из них моторных единиц значительно вариирует. Она больше в т.н. быстрых и меньше в медленных волокнах. При этом возбудимость обоих ниже возбудимости нервных волокон, их иннервирующих. Это зависит от того, что в мышцах разница Е0-Е к больше, и, значит, реобаза выше. ПД достигает 110-130 мв, длительность его 3-6 мсек. Максимальная частота быстрых волокон - около 500 в сек., большинства скелетных - 200-250 в сек. Длительность ПД в медленных волокнах примерно в 2 раза больше, продолжительность волны сокращения - в 5 раз больше, а скорость ее проведения в 2 раза медленнее. Кроме того, быстрые волокна делятся в зависимости от скорости сокращения и лабильности на фазные и тонические.
Скелетные мышцы в большинстве случаев являются смешанными: они состоят как из быстрых, так и медленных волокон. Но в пределах одной моторной единицы все волокна всегда одинаковы. Поэтому и моторные единицы делят на быстрые и медленные, фазные и тонические. Смешанный тип мышцы позволяет нервным центрам использовать одну и ту же мышцу как для осуществления быстрых, фазных движений, так и для поддержания тонического напряжения.
Существуют, однако, мышцы, состоящие преимущественно из быстрых или из медленных моторных единиц. Такие мышцы часто тоже называются быстрыми (белыми) и медленными (красными). Длительность волны сокращения наиболее быстрой мышцы - внутренней прямой мышцы глаза - составляет всего 7,5 мсек., у медленной камбаловидной - 75 мсек. Функциональное значение указанных различий становится очевидным при рассмотрении их ответов на ритмические стимулы. Для получения гладкого тетануса медленной мышцы достаточно раздражать ее с частотой 13 стимулов в сек. в быстрых же мышцах гладкий тетанус возникает при частоте 50 стимулов в сек. В тонических моторных единицах длительность сокращения на одиночный стимул может достигать 1 секунды.
Суммация сокращений моторных единиц в целой мышце . В отличие от мышечных волокон в моторной единице, которые синхронно, одновременно возбуждаются в ответ на приходящий импульс, мышечные волокна различных моторных единиц в целой мышце работают асинхронно. Объясняется это тем, что разные моторные единицы иннервируются различными двигательными нейронами, которые посылают импульсы с различной частотой и разновременно. Несмотря на это суммарное сокращение мышцы в целом имеет в условиях нормальной деятельности слитный характер. Это происходит потому, что соседняя моторная единица (или единицы) всегда успевают сократиться раньше, чем успевают расслабиться те, которые уже возбуждены. Сила мышечного сокращения зависит от числа моторных единиц, вовлеченных одновременно в реакцию, и от частоты возбуждения каждой из них.
Тонус скелетных мышц . В покое, вне работы, мышцы в организме не являются
полностью расслабленными, а сохраняют некоторое напряжение, называемое тонусом. Внешним выражением тонуса является определенная упругость мышц.
Электрофизиологические исследования показывают, что тонус связан с поступлением к мышце редких нервных импульсов, возбуждающих попеременно различные мышечные волокна. Эти импульсы возникают в мотонейронах спинного мозга, активность которых, в свою очередь поддерживается импульсами, исходящими из как из вышестоящих центров, так и из проприорецепторов (мышечных веретен и др.), находящихся в самих мышцах. О рефлекторной природе тонуса скелетных мышц свидетельствует тот факт, что перерезка задних корешков, по которым чувствительные импульсы от мышечных веретен поступают в спинной мозг, приводит к полному расслаблению мышцы.
Работа и сила мышц . Величина сокращения (степень укорочения) мышцы при данной силе раздражения зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект, при сильном растяжении сокращенные мышцы расслабляются. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения падает.
Для измерения силы мышцы определяют либо тот максимальный груз, который она в состоянии поднять, либо максимальное напряжение, которое она может развить в условиях изометрического сокращения. Эта сила может быть очень велика. Так, установлено, что собака мышцами челюсти может поднять груз, превышающий вес ее тела в 8,3 раза.
Одиночное мышечное волокно может развивать напряжение, достигающее 100-200 мг. Учитывая, что общее число мышечных волокон в теле человека равно приблизительно 15-30 млн., они могли бы развить напряжение в 20-30 тонн, если бы все они одновременно тянули в одну сторону.
Сила мышц при прочих равных условиях зависит от ее поперечного сечения. Чем больше сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. При этом имеется ввиду т.н. физиологическое поперечное сечение, когда линия сечения идет перпендикулярно мышечным волокнам, а не мышце в целом. Сила мышц с косыми волокнами больше, чем с прямыми, так как физиологическое ее сечение больше при одинаковом геометрическом. Чтобы сравнить силу разных мышц, максимальный груз (абсолютная сила мышцы), который мышца в состоянии поднять, делят на площадь физиологического поперечного сечения (кг/см.кв.) Таким образом вычисляют удельную абсолютную силу мышцы. Для икроножной мышцы человека она равна 5,9 кг/см.кв., сгибателя плеча - 8,1 кг/см.кв., трехглавой мышцы плеча - 16,8 кг/см.кв..
Работа мышц измеряется произведением поднятого груза на величину укорочения мышцы. Между грузом, который поднимает мышца, и выполняемой ею работой существует следующая закономерность. Внешняя работа мышцы равна нулю, если мышца сокращается без нагрузки. По мере увеличения груза работа сначала увеличивается, а затем постепенно падает. Наибольшую работу мышца совершает при некоторых средних нагрузках. Поэтому зависимость работы и мощности от нагрузки получила название правила (закона) средних нагрузок .
Работа мышц, при которой происходит перемещение груза и движение костей в суставах, называется динамической. Работа мышцы, при которой мышечные волокна развивают напряжение, но почти не укорачиваются - статической. Пример - вис на шесте. Статическая работа более утомительна, чем динамическая.
Утомление мышцы . Утомлением называется временное понижение работоспособ-
ности клетки, органа или целого организма, наступающее в результате работы и исчезающее после отдыха.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен небольшой груз, то амплитуда ее сокращений постепенно убывает, пока не сойдет до нуля. Регистрируется кривая утомления. Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения, удлиняется период расслабления мышцы и увеличивается порог раздражения, т.е. понижается возбудимость. Все эти изменения возникают не сразу после начала работы, существует некоторый период, в течение которого наблюдается увеличение амплитуды сокращений и небольшое повышение возбудимости мышцы. При этом она становится легко растяжимой. В таких случаях говорят, что мышца "врабатывается", т.е. приспосабливается к работе в заданном ритме и силе раздражения. После периода врабатываемости наступает период устойчивой работоспособности. При дальнейшем длительном раздражении наступает утомление мышечных волокон.
Понижение работоспособности изолированной из организма мышцы при ее длительном раздражении обусловлено двумя основными причинами. Первой из них является то, что во время сокращений в мышце накапливаются продукты обмена веществ (фосфорная кислота, связывающая Са++, молочная кислота и др.), оказывающие угнетающее действие на работоспособность мышцы. Часть этих продуктов, а также ионы Са диффундируют из волокон наружу в околоклеточное пространство и оказывают угнетающее действие на способность возбудимой мембраны генерировать ПД. Так, если изолированную мышцу, помещенную в небольшой объем жидкости Рингера, довести до полного утомления, то достаточно только сменить омывающий ее раствор, чтобы восстановились сокращения мышцы.
Другой причиной развития утомления изолированной мышцы является постепенное истощение в ней энергетических запасов. При длительной работе резко уменьшается содержание в мышце гликогена, вследствие чего нарушаются процессы ресинтеза АТФ и КФ, необходимых для осуществления сокращения.
Следует оговорить, что в естественных условиях существования организма утомление двигательного аппарата при длительной работе развивается совершенно не так, как в эксперименте с изолированной мышцей. Обусловлено это не только тем, что в организме мышца непрерывно снабжается кровью, и, следовательно, получает с ней необходимые питательные вещества и освобождается от продуктов обмена. Главное отличие состоит в том, что в организме возбуждающие импульсы приходят к мышце с нерва. Нервно-мышечный синапс утомляется значительно раньше, чем мышечное волокно, в связи с быстрым истощением запасов наработанного медиатора. Это вызывает блокаду передачи возбуждений с нерва на мышцу, что предохраняет мышцу от истощения, вызываемого длительной работой. В целостном же организме еще раньше утомляются при работе нервные центры, (нервно-нервные контакты).
Роль нервной системы в утомлении целостного организма доказывается исследованиями утомления в гипнозе (гиря-корзина), установлением влияния на утомления "активного отдыха", роли симпатической нервной системы (феномен Орбели-Гинецинского) и др..
Для изучения мышечного утомления у человека пользуются эргографией. Форма кривой утомления и величина произведенной работы чрезвычайно вариирует у разных лиц и даже у одного и того же исследуемого при различных условиях.
Рабочая гипертрофия мышц и атрофия от бездействия . Систематическая интенсивная работа мышцы приводит к увеличению массы мышечной ткани. Это явление названо рабочей гипертрофией мышцы. В ее основе лежит увеличение массы протоплазмы мышечных волокон и числа содержащихся в них миофибрилл, что приводит к увеличению диаметра каждого волокна. При этом в мышце происходит активация синтеза нуклеиновых кислот и белков и увеличивается содержание АТФ и КФЫ, а также гликогена. В результате сила и скорость сокращения гипертрофированной мышцы возрастают.
Увеличению числа миофибрилл при гипертрофии способствует преимущественно статическая работа, требующая большого напряжения (силовая нагрузка). Даже кратковременные упражнения, проводимые ежедневно в условиях изометрического режима, достаточны для того, чтобы произошло увеличение числа миофибрилл. Динамическая мышечная работа, производимая без особых усилий, не приводит к гипертрофии мышцы, но может оказывать влияние на весь организм в целом, повышая устойчивость его к неблагоприятным факторам.
Противоположным рабочей гипертрофии явлением служит атрофия мышц от бездействия. Она развивается во всех случаях, когда мышцы почему-то утрачивают способность совершать свою нормальную работу. Это происходит, например, при длительном обездвиживании конечности в гипсовой повязке, долгом пребывании больного в постели, перерезке сухожилия и т.п. При атрофии мышц диаметр мышечных волокон и содержание в них сократительных белков, гликогена, АТФ и других важных для сократительной деятельности веществ резко уменьшается. При возобновлении нормальной работы мышцы атрофия постепенно исчезает. Особый вид мышечной атрофии наблюдается при денервации мышцы, т.е. после перерезки ее двигательного нерва.
Гладкие мышцы Функции гладких мышц в разных органах .
Гладкая мускулатура в организме находится во внутренних органах, сосудах, коже. Гладкие мышцы способны осуществлять относительно медленные движения и длительные тонические сокращения.
Относительно медленные, часто ритмические сокращения гладких мышц стенок полых органов (желудка, кишок, протоков пищеварительных желез, мочеточников, мочевого пузыря, желчного пузыря и т.д.) обеспечивают перемещение содержимого. Длительные тонические сокращения гладких мышц особенно резко выражены в сфинктерах полых органов; их сокращение препятствует выходу содержимого.
В состоянии постоянного тонического сокращения находятся также гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол. Тонус мышечного слоя стенок артерий регулирует величину их просвета и тем самым уровень кровяного давления и кровоснабжения органов. Тонус и двигательная функция гладких мышц регулируется импульсами, поступающими по вегетативным нервам, гуморальными влияниями.
Физиологические особенности гладких мышц. Важным свойством гладкой мышцы является ее большая пластичность , т.е. способность сохранять приданную растяжением длину без изменения напряжения. Скелетная мышца, наоборот, сразу укорачивается после снятия груза. Гладкая мышца остается растянутой до тех пор, пока под влиянием какого-либо раздражения не возникает ее активного сокращения. Свойство пластичности имеет большое значение для нормальной деятельности полых органов - благодаря ему давление внутри полого органа относительно мало изменяется при разной степени его наполнения.
Существуют различные типы гладких мышц. В стенках большинства полых органов находятся мышечные волокна длиной 50-200 мк и диаметром 4-8 мк, которые очень тесно примыкают друг к другу, и потому при рассмотрении их в микроскоп создается впечатление, что они морфологически составляют одно целое. Электронно-микроскопическое исследование показывает, однако, что они отделены друг от друга межклеточными щелями, ширина которых может быть равна 600-1500 ангстрем. Несмотря на это, гладкая мышца функционирует как одно целое. Это выражается в том, что ПД и медленные волны деполяризации беспрепятственно распространяются с одного волокна на другое.
В некоторых гладких мышцах, например, в ресничной мышце глаза, или мышцах радужной оболочки, волокна расположены раздельно, и каждое имеет свою иннервацию. У большинства же гладких мышц двигательные нервные волокна расположены только на небольшом числе волокон.
Потенциал покоя гладкомышечных волокон, обладающих автоматией, обнаруживает постоянные небольшие колебания. Величина его при внутриклеточном отведении равна 30-70 мв. Потенциал покоя гладкомышечных волокон, не обладающих автоматией, стабилен и равен 60-70 мв. В обоих случаях его величина меньше потенциала покоя скелетной мышцы. Это связано с тем, что мембрана гладкомышечных волокон в покое характеризуется относительно высокой проницаемостью для ионов Na. Потенциалы действия в гладких мышцах также несколько ниже, чем в скелетных. Превышение над потенциалом покоя - не больше 10-20 мв.
Ионный механизм возникновения ПД в гладких мышцах несколько отличается от имеющегося в скелетных. Установлено, что регенеративная деполяризация мембраны, лежащая в основе потенциала действия в ряде гладких мышц, связана с повышением проницаемости мембраны для ионов Са++, а не Na+.
Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно медленное снижение мембранного потенциала покоя, которое при достижении определенного уровня сопровождается возникновением ПД.
Проведение возбуждения по гладкой мышце . В нервных и скелетных мышечных волокнах возбуждение распространяется посредством локальных электрических токов, возникающих между деполяризованным и соседними покоящимися участками клеточной мембраны. Этот же механизм свойственен и гладким мышцам. Однако, в отличие от того, что имеет место в скелетных мышцах, в гладких потенциал действия, возникающий в одном волокне, может распространяться на соседние волокна. Обусловлено это тем, что в мембране гладкомышечных клеток в области контактов с соседними имеются участки относительно малого сопротивления, через которые петли тока, возникшие в одном волокне, легко переходят на соседние, вызывая деполяризацию их мембран. В этом отношении гладкая мышца сходна с сердечной. Отличие заключается только в том, что в сердце от одной клетки возбуждается вся мышца, а в гладких мышцах ПД, возникший в одном участке, распространяется от него лишь на определенное расстояние, которое зависит от силы приложенного стимула.
Другая существенная особенность гладких мышц заключается в том, что распространяющийся ПД возникает в низ только в том случае, если приложенный стимул возбуждает одновременно некоторое минимальное число мышечных клеток. Эта "критическая зона" имеет диаметр около 100 мк, что соответствует 20-30 параллельно лежащим клеткам. Скорость проведения возбуждения в различных гладких мышцах составляет от 2 до 15 см/сек. т.е. значительно меньше, чем в скелетной мышце.
Так же, как и в скелетной мускулатуре, в гладкой потенциалы действия имеют пусковое значение для начала сократительного процесса. Связь между возбуждением и сокращением здесь также осуществляется с помощью Са++. Однако в гладкомышечных волокнах саркоплазматический ретикулюм плохо выражен, поэтому ведущую роль в механизме возникновения сокращения отводят тем ионам Са++, которые проникают внутрь мышечного волокна во время генерации ПД.
При большой силе одиночного раздражения может возникнуть сокращение гладкой мышцы. Латентный период сокращения ее значительно больше, чем скелетной, достигая 0,25-1 сек. Продолжительность самого сокращения тоже велика - до 1 минуты. Особенно медленно протекает расслабление после сокращения. Волна сокращения распространяется по гладкой мускулатуре с той же скоростью, что и волна возбуждения (2-15 см/сек). Но эта медленность сократительной активности сочетается с большой силой сокращения гладкой мышцы. Так, мускулатура желудка птиц способная поднимать 2 кг на 1 кв.мм. своего поперечного сечения.
Вследствие медленности сокращения гладкая мышца даже при редких ритмических раздражениях (10-12 в мин) легко переходит в длительное состояние стойкого сокращения, напоминающее тетанус скелетных мышц. Однако энергетические расходы при таком сокращении очень низки.
Способность к автоматии гладких мышц присуща их мышечным волокнам и регулируется нервными элементами, которые находятся в стенках гладко мышечных органов. Миогенная природа автоматии доказана опытами на полосках мышц кишечной стенки, освобожденных от нервных элементов. На все внешние воздействия гладкая мышца реагирует изменением частоты спонтанной ритмики, следствием чего являются сокращения или расслабления мышцы. Эффект раздражения гладкой мускулатуры кишки зависит от соотношения между частотой стимуляции и собственной частотой спонтанной ритмики: при низком тонусе - редких спонтанных ПД - приложенное раздражение усиливает тонус, при высоком тонусе в ответ на раздражение возникает расслабление, так как чрезмерное учащение импульсации приводит к тому, что каждый следующий импульс попадает в фазу рефрактерности от предыдущего.
Раздражители гладких мышц . Одним из важных физиологически адекватных раздражителей гладких мышц является их быстрое и сильное растяжение. Оно вызывает деполяризацию мембраны мышечного волокна и возникновение распространяющегося ПД. В результате мышца сокращается. Характерной особенностью гладких мышц является их высокая чувствительность к некоторым химическим раздражителям, в частности, к ацетилхолину, норадреналину, адреналину, гистамину, серотонину, простагландинам. Эффекты, вызываемые одним и тем же химическим агентом, в разных мышцах и при различном их состоянии могут быть неодинаковы. Так, АХ возбуждает гладкие мышцы большинства органов, но тормозит мышцы сосудов. Адреналин расслабляет небеременную матку, но сокращает беременную. Эти различия связаны с тем, что указанные агенты реагируют на мембране с различными химическим рецепторами (холино-рецепторами, альфа и бета адренорецепторами), и в итоге по разному изменяют ионную проницаемость и мембранный потенциал гладкомышечных клеток. В тех случаях, когда раздражающий агент вызывает деполяризацию мембраны, возникает возбуждение, и, наоборот, гиперполяризация мембраны под влиянием химического агента приводит к торможению активности и расслаблению гладкой мышцы.